
Charnia, evtl. eine Art Federkoralle.
Quelle: Wikipedia File:Charnia.png, dort lizensiert unter der Creative Commons Attribution 2.5 License.
Nachdem die letzte der globalen Eiszeiten vor etwa 580 Millionen Jahren zuende gegangen war, entstand auf der Erde wieder ein Klima, in dem sich Leben entwickeln und ausbreiten konnte. Die Zeit der letzten Vereisungen und der anschließenden freundlicheren Phase vor 630 bis vor 542 Millionen Jahren bezeichnet man als Ediacarium (früher auch als Vendium bezeichnet). Streng genommen gehört das Ediacarium noch zur Zeit vor dem Erdaltertum, die man auch Präkambrium oder neuerdings Proterozoikum nennt. Ich habe diese Zeit dennoch hier mit hinzugenommen, da sich im späten Ediacarium nach dem Ende der großen Vereisungen vielzelliges Leben im Meer auszubreiten begann. Diese Entwicklung hat sich im Kambrium vor 542 bis 488 Millionen Jahren dann fortgesetzt und beschleunigt.
Nach dem Ende der globalen Eiszeiten muss sich eine große Zahl neuer Lebensräume im Ozean aufgetan haben, deren Besetzung erstmalig in der Erdgeschichte die Entstehung vieler verschiedener mehrzelliger Tier- und Pflanzenarten zur Folge hatte. Diese Lebewesen besaßen allerdings noch keine Kalkschalen oder Kalkskelette, sodass sie kaum fossile Spuren hinterließen. Erst nach dem Ende des zweiten Weltkrieges sind Fossilien aus dieser Zeit entdeckt und auch richtig interpretiert worden, insbesondere in den Ediacara-Hügeln in den Flinders Ranges in Süd-Australien -- daher auch der Name des Zeitalters, der erst im Mai 2004 offiziell beschlossen wurde. Man spricht auch von der Ediacara-Fauna. Hier einige Lebewesen aus dieser Zeit:



Vor 542 Millionen Jahren, beim Übergang vom Ediacarium zum Kambrium, starb ein großer Teil dieser Lebewesen aus, und neue Lebensformen verbreiteten sich. Das Land war weiterhin unbewohnt und unbewachsen, aber im Meer begann die sogenannte Kambrische Explosion, in der innerhalb von nur 50 Millionen Jahren alle modernen Tierstämme entstanden.
Zu Beginn des Kambriums gab es zum ersten Mal Weichtiere mit Schalen oder anderen festen Bestandteilen in großer Zahl, was das Finden entsprechender Fossilien sehr erleichtert. Man nennt diese Tierwelt daher auch small shelly fauna (also kleine schalige Tierwelt, siehe Wikipedia: Small shelly fauna ). Die Fossilien aus dieser Zeit sind tatsächlich meist nur wenige Millimeter groß. Allerdings lassen sich diese eigenartigen hartschaligen Winzlinge nur schwer den heute bekannten Tiergruppen zuordnen. Eine Tiergruppe aus dieser Zeit gibt es allerdings heute noch: die Armfüßer (Brachiopoden), die ähnlich wie Muscheln aussehen, aber eine zweiklappige Schale besitzen. Armfüßer sind gleichsam die Muscheln des Erdaltertums, sind aber anders als Muscheln aufgebaut (zu Armfüßern und Muscheln siehe auch Kapitel 4.6 Perm).

Eine andere Tiergruppe aus dem Beginn des Kambriums sind die Archaeocyathiden.
Diese Tiergruppe hat Ähnlichkeit mit den heutigen Schwämmen und Korallen.
Wie diese bildeten sie erste flache Riffe im Meer.
Allerdings starben sie bereits vor 513 Millionen Jahren wieder aus -- warum, ist unbekannt.
Etwa vor 530 Millionen Jahren entwickelten sich die Arthropoden (Gliederfüßer) zur vorherrschenden Tiergruppe in den Meeren des Kambriums. Besonders bekannt sind die Trilobiten (siehe auch http://www.trilobites.info/ ), die erst am Ende des Perm vor circa 250 Millionen Jahren beim größten globalen Massensterben ausstarben. Damit haben die Trilobiten die Ozeane also über fast 200 Millionen Jahre hinweg mit einer riesigen Artenvielfalt bevölkert. Heute gehören zu den Gliederfüßern so unterschiedliche Tiergruppen wie Insekten, Tausendfüßer, Krebse, Entenmuscheln, Spinnen, Skorpione und Milben.




Links: Wiwaxia wurde 1 bis 5 cm groß.
Die Zuordnung zu heutigen Tiergruppen ist unklar.
Sie könnte zu den Weichtieren oder zu den Ringelwürmern gehören, zu denen
auch der Regenwurm gehört.
Quelle:
Wikimedia Commons File:Wiwaxia corrugata.jpg,
Autor nicht erkennbar,
dort lizensiert unter der
Creative Commons Attribution 2.5 License.
Rechts:
Pikaia: dieses etwa 5 cm lange fischförmige Wesen war ein frühes Wirbeltier.
Quelle:
Wikimedia Commons File:Pikaia BW.jpg von Nobu Tamura,
dort lizenziert unter der Creative Commons–Lizenz „Attribution 3.0 Unported“.
Das Erdklima wurde während des Kambriums immer wärmer, die Polkappen schmolzen und der Meeresspiegel stieg stark an. Vor etwa 513 Millionen Jahren erreichte der Meeresspiegel einen Höchststand, und große Teile der Kontinente waren von flachen Meeren überflutet. Am Südpol lag ein großer Kontinent, Gondwana genannt. Drei kleinere Kontinente, die heute Nordamerika, Nordosteuropa und Sibirien entsprechen, befanden sich wie größere Inseln in Äquatornähe. Mitteleuropa existierte noch nicht -- seine Teile lagen alle in südlichen Breiten. Das Klima war heiß und feucht, die Pole waren das ganze Jahr hindurch eisfrei, und im Inneren der Kontinente erstreckten sich ausgedehnte Wüsten.


Es gibt aus dieser Zeit (mittleres Kambrium) eine berühmte Fundstätte für Fossilien:
der Burgess Shale (Burgess-Schiefer)
im Yoho National Park in den Rocky Mountains, nahe der Stadt Field in British Columbia (Kanada).
Vor 500 Millionen Jahren erschienen erste Korallen (die sogenannten tabulaten Korallen, die am Ende des Perm vor circa 250 Millionen Jahren beim großen globalen Massensterben wieder ausstarben), und erste Schnecken (Gastropoden). Heute gehören 78% aller bekannten Weichtiere zu den Schnecken. Auch erste Kopffüßer (Cephalopoden) erschienen -- zu ihnen gehören die Gruppen der Perlboote (Nautiloideen), Ammoniten, Bactriten und die Tintenfische (Coleoidea). Insbesondere die Perlboote (Nautiloideen) verbreiteten sich zum Ende des Kambriums stark.


Links: Ammoniten,
Quelle: Wikipedia Commons,
Quelle dort:
The Wonderful Paleo Art of Heinrich Harder,
Autor: Heinrich Harder (1858-1935),
Public Domain (copyright abgelaufen).
Rechts: Perlboote (Nautiloideen) sind lebende Fossilien.
Quelle: Wikipedia Commons,
demnach freigegeben (public domain) durch den Autor TheMadBaron im English Wikipedia project.
Vor 488 Millionen Jahren schließlich endete das Kambrium mit einem Massensterben, bei dem
viele der vorherrschenden Arten ausstarben und sich die Tier- und Pflanzenwelt der Ozeane grundlegend
änderte. Der Grund dafür ist unklar -- vielleicht sind neuerlich Vereisungen dafür verantwortlich zu machen.
a) Die Entstehung von Zähnen und Knochen
b) Chordatiere, Lanzettfischchen und Kiemenbögen
c) Die Evolution der Augen
Bei Geologen sind die sogenannten Conodonten-Elemente sehr bekannt: winzige, meist etwa 0,2 bis 0,5 mm große sehr vielgestaltige harte Mikrofossilien, die man in großer Zahl in den versteinerten Sedimenten urzeitlicher Meere findet. Ihr Alter liegt zwischen 570 und 225 Millionen Jahren, d.h. sie sind vom Kambrium bis zum Massensterben am Ende der Trias zu finden. Neil Shubin schreibt in seinem Buch Der Fisch in uns auf S. 95:
... hat man Conodonten(-Elemente) auf allen Kontinenten entdeckt, und an manchen Stellen kann man keinen Stein aufschlagen, ohne sie in großer Zahl zu finden.
Übrigens hat man früher zur Verwirrung vieler Nicht-Fachleute die Conodonten-Elemente oft auch einfach als Conodonten bezeichnet, während man heute das Tier so bezeichnet, von dem sie stammen (siehe unten).
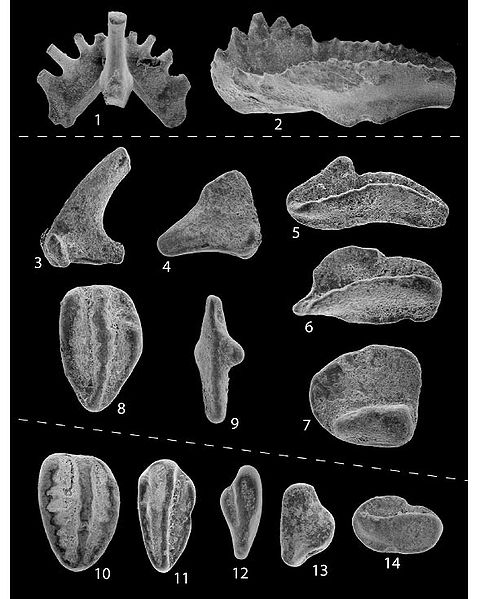
Für Geologen sind die Conodonten-Elemente sehr nützlich, denn
sie bilden hervorragende Leitfossilien zur Altersbestimmung von Gesteinen,
da sie so häufig vorkommen und ihre jeweiligen Ausprägungen
charakteristisch für einzelne Zeitabschnitte sind.
Dabei blieb lange unklar, was Conodonten-Elemente eigentlich genau sind --
Teile von Muschelschalen, Schnecken, Gliederwürmern, Wirbeltieren oder was sonst?
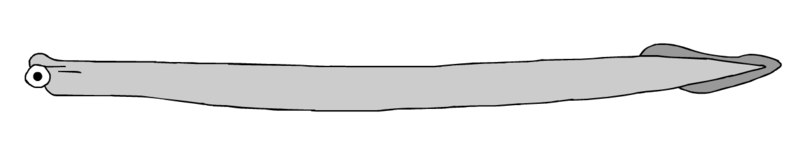
Das Rätsel wurde erst im Jahr 1982 zufällig gelöst, als der britische Forscher Clarkson sich eine Sammlung von Gesteinsplatten aus dem Unter-Karbon ansah, die das Britische Geologische Amt Edinburgh bereits seit dem Jahr 1925 aufbewahrt hatte. Dabei fand er ein etwa 4 cm langes und 2 mm breites wurmähnliches Tier mit den berühmten Conodonten-Elementen im Kopfbereich. Das erste Conodonten-Tier war gefunden (Bild siehe oben)!
Damit war klar: die Conodonten-Elemente sind Zahnstrukturen urtümlicher Fische (Conodonten). Sie sind damit die ältesten fossilen Zähne in der Natur. Die Conodonten selbst sind weiche aalähnliche Tiere ohne harte Knochen, Schädel oder Kiefer. Vermutlich handelt es sich bei ihnen um Stammformen der Wirbeltiere. Eine Chorda Dorsalis besitzen sie bereits, d.h. sie gehören auf jeden Fall wie das Lanzettfischchen zu den Chordatieren, haben aber im Gegensatz zum Lanzettfischchen bereits Zahnstrukturen (Lanzettfischchen sind dagegen ähnlich wie Seescheiden zahnlose Filtrierer -- mehr dazu siehe unten).
Mit den harten Zahnstrukturen waren Conodonten in der Lage, anderen Fischen als Räuber gefährlich zu werden. Ein Wettrennen setzte ein, bei dem die Räuber zusätzlich Kiefer entwickelten, während die potentiellen Beutefische sich Panzer zulegten. So entstanden die sogenannten Ostracodermi (Knochenhäuter): kieferlose Fische, die im Kopfbereich einen Schädel mit einem Knochenpanzer besaßen. Betrachtet man diesen Knochenpanzer unter dem Mikroskop, so erkennt man, dass er aus winzigen miteinander verschmolzenen Zähnchen besteht. Neil Shubin schreibt dazu in seinem Buch Der Fisch in uns auf S. 99: Zähne entstanden ursprünglich, um andere Tiere zu beißen; später dienten sie in veränderter Form als Schutz.
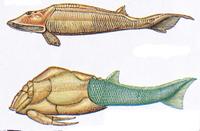
Das Material, das Zähne und Knochen bei Wirbeltieren so hart macht, ist das Mineral Hydroxylapatit,
eine speziellen Form von Kalziumphosphat. Knochen bestehen zu etwa 40 %, Zahnschmelz sogar zu 95 % aus diesem Mineral.
Das harte Gewebe anderer Tiere wie Muscheln, Krebse oder Insekten enthält dieses Mineral dagegen nicht;
bei ihnen kommen statt dessen andere Stoffe wie Kalziumkarbonat oder Chitin zum Einsatz.
Aufgrund der Dauerhaftigkeit von Hydroxylapatit sind Zähne meist die am besten erhaltenenen Überreste von Lebewesen,
wie die Conodonten-Elemente eindrucksvoll belegen.
Letztlich entwickelten sich die Zähne als spezielle Strukturen der Haut und sind daher eng mit deren Evolution verknüpft. Die winzigen Hautzähnchen auf der Haut von Haien macht diesen engen Zusammenhang auch heute noch deutlich. Ganz ähnlich wie Zähne haben sich später auch andere Körperelemente aus Hautstrukturen entwickelt: Schuppen, Haare, Federn, Schweißdrüsen und Milchdrüsen.
Die Chordatiere (Chordata) umfassen die Wirbeltiere und ihre Verwandten, die an Stelle einer Wirbelsäule zumindest über eine Art Stützstab im Rücken verfügen -- die Chorda dorsalis (Notochord), nach der die Chordatiere auch benannt sind und die man als Vorläufer der Wirbelsäule ansehen kann.
Die entwicklungsgeschichtlich ältesten Chordatiere sind vermutlich die Manteltiere, zu denen die Seescheiden gehören (siehe Buch). Hinzu kommt die Gruppe der Schädellosen sowie als entwicklungsgeschichtlich jüngste Gruppe die Wirbeltiere, die über einen Schädel und eine Wirbelsäule verfügen. Noch vor Schädel und Wirbelsäule sind die Zähne entstanden, wie die Conodonten zeigen (siehe oben).
Das bekannteste urtümliche heute noch lebende Chordatier ist neben der Seescheide sicher das Lanzettfischchen, das trotz seines Namens noch kein Fisch ist, sondern eher einem Wurm gleicht. Es besitzt noch keinen Schädel, keine Wirbelsäule, keine Extremitäten (Flossen) und auch noch keine Zähne. Auch Augen sucht man vergeblich -- immerhin gibt es aber sogenannte Rückenmarksaugen im Neuralrohr, also kleine Lichtsinnesorgane, die Licht wahrnehmen können, auch wenn das mit echtem Sehen noch wenig gemeinsam hat.
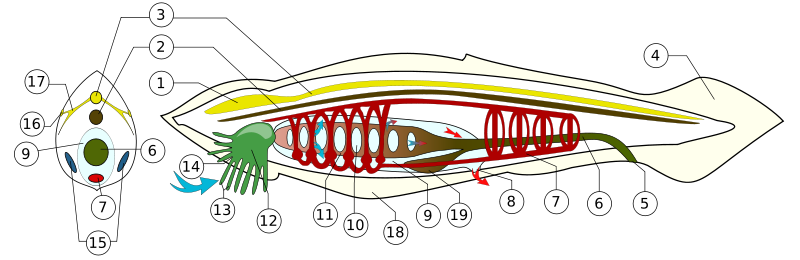
Lanzettfischchen sind wie Seescheiden Filtrierer und verfügen wie diese über einen
sogenannten Kiemendarm, der sich aus dem vorderen Bereich des Darms entwickelt hat.
Mit diesem filtern sie Nahrungsteilchen aus dem Wasser und nehmen zugleich Sauerstoff auf,
wobei die Sauerstoffaufnahme eher ein Nebeneffekt ist, da ein großer Teil des Sauerstoffs
auch direkt über die Haut aufgenommen wird.
Kiemen haben sich also zunächst als Nahrungsfilter entwickelt und wurden erst später bei den Fischen
zu reinen Atmungsorganen umfunktioniert.
Das Wasser verlässt bei den Lanzettfischchen den Kiemendarm über Kiemenspalten, die sich zwischen sogenannten Kiemenbögen befinden. Diese Kiemenbögen bilden letztlich die Basis für die Entwicklung der verschiedenen Kopfstrukturen bei den Wirbeltieren. Neil Shubin hat in seinem Buch Der Fisch in uns in Kapitel 5 eindrucksvoll beschrieben, wie sich der Ursprung verschiedener Bereiche unseres Kopfes auf einzelne Kiemenbögen zurückführen lässt, die wir im Embryonalstadium noch besitzen. Hier einige Details dazu:
Unser Kopf ist anatomisch zunächst recht verwirrend aufgebaut. Manche Nerven und Blutgefäße nehmen beispielsweise sehr umständliche Wege. Das liegt daran, dass unser Kopf eine lange Entwicklungsgeschichte besitzt, die ihren Anfang in den Kiemenbögen nimmt und die sich in der Entwicklung des Embyos widerspiegelt.
Nach gut 3 Wochen besitzen menschliche Embryos (und ebenso die Embryos der meisten Wirbeltiere) 4 winzige Kiemenbögen, aus denen sich nach und nach die unterschiedlichen Kopfstrukturen entwickeln, wobei aus jedem Kiemenbogen eine miteinander verbundene Struktur aus Knochen, Muskeln, Blutgefäßen und Nerven entsteht (genau genommen gibt es beim Menschen 2 weitere Kiemenbögen, die aber nur wenig Bedeutung tragen, sodass wir sie hier weglassen).
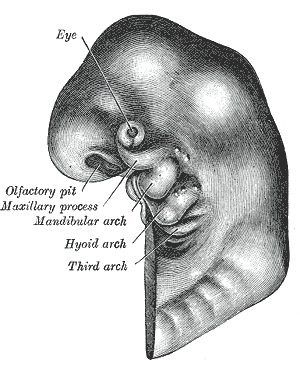
Bei Haien, die entwicklungsgeschichtlich zu den recht urtümlichen Fischen gehören,
sind die aus den Kiemenbögen entstehenden Strukturen noch einigermaßen übersichtlich,
während bei uns Menschen das Heranwachsen des Kopfes über mehrere Umbauphasen
schließlich zu einem recht komplexen Gebilde führt, bei dem man die ursprüngliche Segmentstruktur kaum
noch wiedererkennt, auch wenn sie grundsätzlich noch vorhanden ist.
Hier eine kurze Übersicht, welcher Kiemenbogen welche Strukturen bei uns Menschen hervorbringt:
Besonders interessant ist dabei, dass das Innenohr nicht einheitlich aus einem einzigen Kiemenbogen entsteht, sondern aus zweien. Das liegt daran, dass unsere Gehörstrukturen mit den drei Gehörknöchelchen Hammer, Amboss und Steigbügel in dieser Form erst mit der Entwicklung der Säugetiere entstanden sind, wobei bereits vorhandene Grundstrukturen (beispielsweise Kieferknochen) zweckentfremdet wurden und eine neue Aufgabe erhalten haben (mehr dazu an anderer Stelle).
Es gibt eine große Vielfalt verschiedener Augentypen im Tierreich, die in ihrer jeweiligen Bauweise meist unabhängig voneinander entstanden sind, da sich die entsprechenden Abstammungslinien bereits vor der Entstehung dieser Augen voneinander getrennt hatten. Dabei können mehrfach recht ähnliche Augentypen entstehen wie beispielsweise die Linsenaugen bei Wirbeltieren und Tintenfischen, aber auch ganz andere Bauformen wie das Facettenauge bei Insekten. Die Fähigkeit, Licht wahrnehmen zu können, bringt offenbar so große Vorteile, dass die Evolution immer wieder neue Wege findet, diese Vorteile zu nutzen. Man schätzt, dass auf diese Weise Augen rund 40 bis 50 Mal unabhängig voneinander neu entstanden sind.
Die biochemischen Grundlagen für die Wahrnehmung von Licht dürften schon vor sehr langer Zeit bei Einzellern entstanden sein. Sie bilden eine gemeinsame Basis für die Lichtwahrnehmung bei allen vielzelligen Tieren. Das Schlüsselmolekül ist dabei das Protein Opsin in seinen verschiedenen Varianten, das zusammen mit verschiedenen Vitamin-A-Abkömmlingen ein lichtempfindliches Molekül bildet: das Sehpigment. Licht lässt dieses Molekül wieder in seine beiden Teile zerbrechen, woraufhin das freiwerdende Opsin den Transport chemischer Stoffe durch die Zellmembran beeinflusst und so eine Signalkette auslöst. Es dauert anschließend einige Minuten, bis sich die beiden Teile wieder zum lichtempfindlichen Sehpigment zusammenfinden. Wenn wir aus dem hellen Sonnenlicht in einen dunklen Raum treten, können wir daher zunächst darin kaum etwas erkennen, bis sich wieder genügend viele Sehpigment-Moleküle regeneriert haben.
Auch der genetische Werkzeugkasten zur Entstehung von Augen hat einen gemeinsamen Ursprung. So gibt es ein bestimmtes Gen, das bei fast allen Tieren gleichsam als Hauptschalter zur Entwicklung eines Auges dient (eine Ausnahme ist beispielsweise die Würfelqualle, siehe unten). Bei Fliegen nennt man dieses Gen eyeless (da es zu fehlenden Augen führt, wenn es defekt ist), bei Mäusen dagegen heißt es Pax 6. Dort wo dieses Gen aktiv ist, ensteht ein neues Auge. Man kann sogar das Mäuse-Gen auf ein Fliegenbein bringen und dort aktivieren -- es entsteht ein Auge auf dem Bein, allerdings ein Fliegenauge und kein Mäuseauge (zugegeben -- solche Experimente sind irgendwie gruselig). Ähnlich wie die Hox-Gene für die Strukturierung der Längsachse geben eyeless und Pax 6 also nur eine Ortsinformation an die Zellen und veranlassen sie, ihr individuelles Genprogramm für die Entstehung eines Auges zu starten. Der Hauptschalter für die Bildung eines Auges ist also bereits entstanden, noch bevor sich die Abstammungslinien von Fliegen und Mäusen getrennt haben, während sich die Details im jeweiligen nachgelagerten Augen-Genprogramm erst anschließend entwickelt haben (meist durch Umfunktionieren und Weiterentwicklung bereits vorhandener Gene und Strukturen).
Einen einfachen richtungsabhängigen Lichtsinn findet man schon bei Einzellern wie den Augentierchen (Euglena), die zu den Eukaryoten (Zellen mit Zellkern) gehören. Damit sind sie beispielsweise in der Lage, auf Lichtquellen zuzuschwimmen und sie so besser für die Photosynthese zu nutzen.
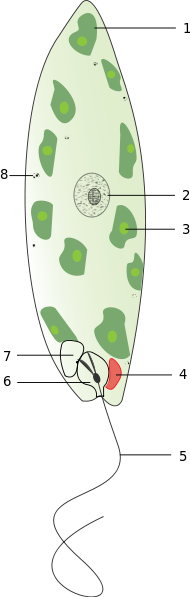
Wirkliche Augen haben sich jedoch wohl erst im Ediacarium und Kambrium entwickelt,
und zwar sowohl bei den Bilateria (Zweiseitentieren) als auch teilweise bei älteren Abstammungslinien
wie beispielsweise den Nesseltieren (Cnidaria),
die sich vor rund einer Milliarde Jahren von unserem Entwicklungszweig abgespalten hatten
(siehe das Beispiel der Würfelqualle weiter unten sowie Kapitel 3.3).
Bei der Aufspaltung der Bilateria in Protostomier (Urmünder, also der Großteil der Wirbellosen wie beispielsweise Weichtiere und Insekten) und Deuterostomier (Neumünder, insbesondere Wirbeltiere) vor rund 600 Millionen Jahren haben sich bei den Lichtsinneszellen zwei verschiedene Grundstrukturen etabliert, mit denen die lichtsammelnde Oberfläche der Lichtsinneszellen jeweils vergrößert wird. Bei den Lichtsinneszellen der Protostomier geschieht dies durch die Ausbildung vieler kleiner Falten (rhabdomere Photorezeptoren), bei den Lichtsinneszellen der Deuterostomier (also u.a. bei uns) wird es dagegen durch viele kleine borstenartige Ausstülpungen erreicht (ziliäre Photorezeptoren). Außerdem reagieren die Lichtsinneszellen bei Protostomiern mit einer Verringerung der elektrischen Spannung auf Lichteinfall, während bei den Deuterostomiern der Lichteinfall die Spannung erhöht (allerdings erst in der Nervenzelle, die mit der Lichtsinneszelle verbunden ist).
Vermutlich haben die beiden verschiedenen Typen von Lichtsinneszellen schon vor der Trennung der Bilateria in Proto- und Deuterostomier existiert. Man hat nämlich ein Tier gefunden, das noch beide Zelltypen (rhabdomere und ziliäre Photorezeptoren) zugleich besitzt. Es handelt sich um einen Borstenwurm (Vielborster oder Polychaeten), und zwar um Platynereis dumerilii (siehe auch http://www.platynereis.de/). Er gilt als lebendes Fossil und dürfte viele Ähnlichkeiten mit den sehr frühen Protostomiern aufweisen. Die einfachen Augen seiner Larven sind mit ihren rhabdomeren Photorezeptoren typisch für Protostomier, aber zugleich besitzen die Larven zusätzlich winzige ziliäre Lichtrezeptoren unter der Haut, die Teil des Nervensystems sind.
Neuere Forschungsergebnisse legen folgendes Szenario nahe: Dort, wo rhabdomere Lichtsinneszellen vorhanden sind, werden nur sie zum Sehen verwendet, während die ziliären Lichtrezeptoren eher der nicht-bildlichen Lichtwahrnehmung dienen, also beispielsweise zur Erfassung von Tag und Nacht. Bei den Deuterostomiern (speziell den Wirbeltieren) haben diese rhabdomeren Lichtsinneszellen dann eine andere Aufgabe übernommen und bilden vermutlich als retinale Ganglienzellen den Sehnerv. Im Gegenzug haben die ziliären Lichtrezeptoren ihr Aufgabenspektrum erweitert und dienen nun nicht mehr nur zur einfachen Lichtwahrnehmung, sondern zum bildlichen Sehen. Mehr dazu siehe in Trevor D.Lamb: Das Auge -- Organ mit Vergangenheit, Spektrum der Wissenschaft, Februar 2012, S. 23.
Der Borstenwurm zeigt auch, dass beide Typen von Lichtsinneszellen einen unterschiedlichen Ursprung haben: Die ziliären Lichtsinneszellen der Deuterostomier stammen von Zellen ab, die dem Nervensystem entstammen (man vergleiche auch das Bild des Lanzettfischchens weiter oben). Im Wirbeltier-Embryo bilden sich die Augen entsprechend als Ausstülpung einer Zellgruppe, aus der auch das Gehirn hervorgeht. Die rhabdomeren Lichtsinneszellen der Protostomier (Weichtiere, Insekten, ...) haben ihren Ursprung dagegen in den Hautzellen, und im Embryo entstehen sie entsprechend aus einer Einstülpung der äußeren Zellschicht, aus der sich auch die Haut bildet. Mehr dazu siehe unter heise online: Wo Darwin noch erschauderte.
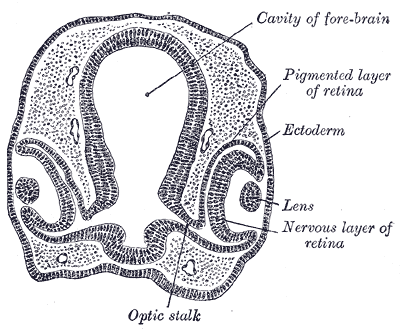
Im Lauf der Zeit ist eine große Vielfalt verschiedener Augentypen entstanden,
die nicht immer auf bestimmte Zweige im Stammmbaum der Tiere beschränkt sind, sondern die
sich oft unabhängig voneinander bei verschiedenen Tiergruppen in unterschiedlicher Detailausführung entwickelt haben.
Hier einige Beispiele:
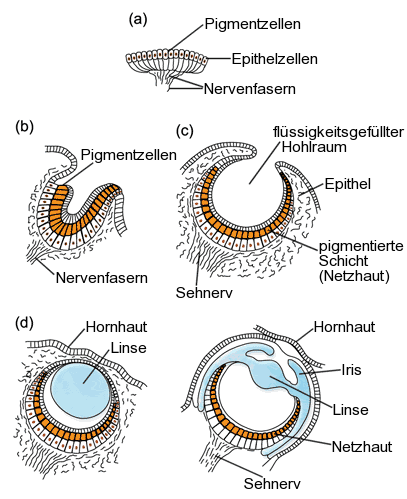
Regenwürmer haben noch keine richtigen Augen, sondern nur einzelne Lichtsinneszellen.
Bei Seesternen und vielen Quallen sind diese Sinneszellen schon in einem bestimmten Areal konzentriert (Flachauge).
Beim Pigmentbecherauge (beispielsweise bei Strudelwürmern, vielen Schnecken und beim Lanzettfischchen) und beim Grubenauge sinkt dieses Areal in Form einer Grube nach innen, sodass sich die Einfallsrichtung des Lichts grob erkennen lässt.
Wenn die Grube sich zu einem Hohlraum mit lochartiger Öffnung weiterentwickelt, haben wir ein sogenanntes Lochauge vor uns. Es funktioniert wie eine Lochkamera und erlaubt bereits das Erkennen von Bildern. Das Lochauge findet man beispielsweise bei Perlbooten (urtümliche Kopffüßer) oder bei bestimmten Meeresschnecken, beispielsweise bei den Seeohren (Haliotidae) und bei den Kreiselschnecken (Trochidae) (siehe http://www.weichtiere.at/Schnecken/index.html?/Schnecken/morphologie/augen.html).

Ein Blasenauge entsteht, wenn die lochartige Öffnung mit einer transparenten Haut verschlossen ist.
Es tritt bei Hohltieren, Schnecken (zum Beispiel der Apfelschnecke Ampullaria) und Ringelwürmern auf.
Krebse und Insekten besitzen ein Facettenauge (Komplexauge), das aus einer großen Anzahl einzelner Augen besteht, die jeweils nur einen kleinen Bildausschnitt erfassen. Die räumliche Auflösung dieser Augen ist relativ mäßig, aber dafür ist ihre zeitliche Auflösung sehr gut -- jeder von uns weiß, wie schnell eine Fliege auf eine herabsausende Hand reagieren kann! Für große Tiere ist ein Facettenauge allerdings ungeeignet -- hier leistet ein Linsenauge bessere Dienste.

Linsenaugen gibt es in verschiedener Komplexität und mit unterschiedlichen Bauplänen.
Es gibt Linsenaugen nicht nur bei Wirbeltieren, sondern unabhängig davon auch
in einfacher Bauweise bei Schnecken (z.B. Weinbergschnecken),
Kopffüßern, bei manchen Gliederfüßern (z.B. Ocellen der Spinnentiere) und sogar bei
den gefürchteten hochgiftigen Würfelquallen (Cubozoa), die noch nicht einmal zu den Bilateria,
sondern zu den Nesseltieren gehören.
So besitzt die Würfelqualle Carybdea marsupialis neben mehreren einfachen schlitzförmigen und grubenförmigen
Pigmentaugen auch mehrere einfache Linsenaugen. Das eigentliche Pax-6-Gen besitzt die Würfelqualle dagegen nicht, d.h.
ihr Entwicklungsweg hatte sich von dem der anderen Tiere bereits getrennt, noch bevor Pax 6 entstanden ist,
das bei den anderen Tieren als Hauptschalter für die Augenentwicklung dient.
Damit ist auch klar, dass die Linsenaugen der Würfelqualle vollkommen unabhängig vom Linsenauge anderer Tiere entstanden sein muss.
Das Augen-Gen der Würfelqualle sieht aus wie eine vereinfachte Mischung der Gene Pax 6 und Pax 2, wobei Pax 2
bei uns für die Entwicklung des Innenohrs wichtig ist.
Bereits im Kambrium vor rund 500 Millionen Jahren besaßen die damaligen Wirbeltiere (urtümliche Fische) komplexe Linsenaugen ähnlich den unseren. Man vermutet, dass weniger als 100 Millionen Jahre für die Entstehung dieser komplexen Augen aus den ersten einfachen Lichtsensoren ausreichten. Unabhängig davon entstanden solche Augen auch bei bestimmten Kopffüßern (Tintenfische, Kraken). Im Embryo bilden sie sich bei Kopffüßern und Wirbeltieren auf vollkommen unterschiedliche Weise: Da Kopffüßer Protostomier sind, entstehen sie bei ihnen als Einstülpung von Hautgewebe, bei Wirbeltieren (die Deuterostomier sind) dagegen als Ausstülpung des Gehirns. Als Folge davon liegen die Lichtsinneszellen in der Netzhaut bei Tintenfischen auf der dem Licht zugewandten Seite, bei Wirbeltieren auf der dem Licht abgewandten Seite (also gleichsam verkehrt herum, was beispielsweise den blinden Fleck zur Folge hat, der bei Tintenfischaugen nicht notwendig ist). Auch der Mechanismus zum Scharfstellen der Linse ist unterschiedlich: Bei Wirbeltieren wird die Linse verformt, bei Tintenfischen wird sie nach vorne oder hinten bewegt. Und ein weiterer Unterschied: die Lichtsinneszellen bei Wirbeltieren und Kopffüßern gehören zu zwei grundverschiedenen Typen, wie wir bereits wissen -- die rhabdomeren Lichtsinneszellen der Kopffüßer sind denen der Insekten ähnlich, da beide zu den Protostomiern gehören, während Wirbeltiere ziliäre Lichtsinneszellen zum bildlichen Sehen verwenden.

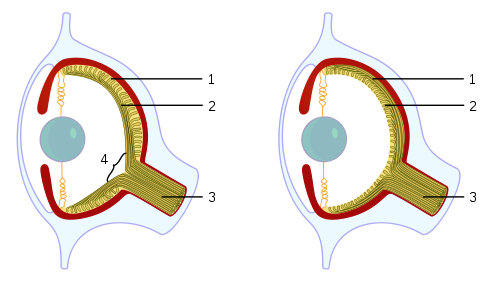
Man sieht: Die Evolution ist keineswegs nur auf einen einzigen Weg angewiesen, sondern es führen sehr
viele Wege zum Ziel, ein funktionsfähiges Auge entstehen zu lassen.
Jeder einzelne dieser Wege mag recht unwahrscheinlich erscheinen und wird womöglich im Detail nur ein einziges Mal beschritten.
Dennoch gibt es genügend Möglichkeiten, sodass Augen nicht nur einmal, sondern viele Dutzend mal
entstehen konnten.
Literatur zu dem Thema:
last modified on 29 June 2012
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}